23 releases
Uses old Rust 2015
| 0.1.22 | Feb 24, 2023 |
|---|---|
| 0.1.20 | Jan 29, 2023 |
| 0.1.18 | Dec 27, 2022 |
| 0.1.17 | Nov 23, 2022 |
| 0.1.0 | Jul 13, 2020 |
#298 in Biology
Used in 4 crates
(3 directly)
500KB
2.5K
SLoC
Quick Probability Inference Engine on RNA Structural Alignment
Installation
This project is written in Rust, a systems programming language. You need to install Rust components, i.e., rustc (the Rust compiler), cargo (the Rust package manager), and the Rust standard library. Visit the Rust website to see more about Rust. You can install Rust components with the following one line:
curl --proto '=https' --tlsv1.2 -sSf https://sh.rustup.rs | sh
Rustup arranges the above installation and enables to switch a compiler in use easily. You can install ConsProb:
# AVX, SSE, and MMX enabled for rustc
# Another example: RUSTFLAGS='--emit asm -C target-feature=+avx2 -C target-feature=+ssse3 -C target-feature=+mmx -C target-feature=+fma'
RUSTFLAGS='--emit asm -C target-feature=+avx -C target-feature=+ssse3 -C target-feature=+mmx' \
cargo install consprob
Check if you have installed ConsProb properly:
# Its available command options will be displayed
consprob
You can run ConsProb with a prepared test set of sampled tRNAs:
git clone https://github.com/heartsh/consprob \
&& cd consprob
cargo test --release
# The below command requires Gnuplot (http://www.gnuplot.info)
# Benchmark results will be found at "./target/criterion/report/index.html"
cargo bench
Advanced Computation of RNA Structural Context Profiles
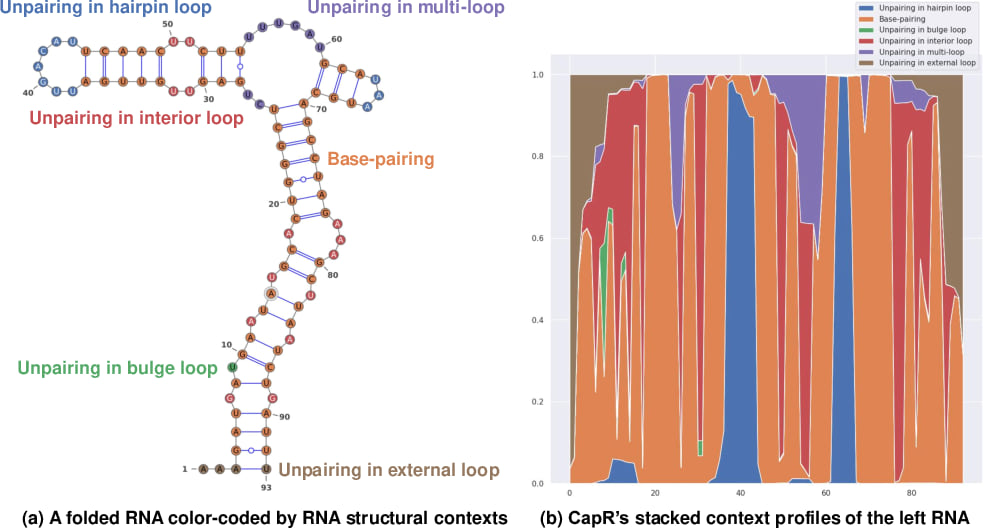
Measuring the structural context profile of each RNA nucleotide (i.e., the posterior probability that each nucleotide is in each structural context type) is beneficial to various structural analyses around functional non-coding RNAs. For example, CapR computes RNA structural context profiles on RNA secondary structures, distinguishing (1) unpairing in hairpin loops, (2) base-pairings, (3) unpairing in bulge loops, (4) unpairing in interior loops, (5) unpairing in multi-loops, and (6) unpairing in external loops as available structural context types:
 |
|---|
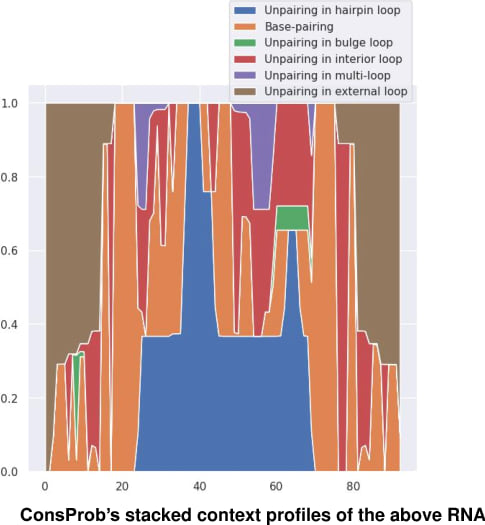
Respecting CapR, ConsProb offers the computation of average structural context profiles on RNA structural alignment, distinguishing the above structural context types. Technically, ConsProb calculates the structural context profile of each nucleotide pair on RNA pairwise structural alignment and averages this pairwise context profile over available RNA homologs to each RNA homolog, marginalizing these available RNA homologs. ConsProb's context profile computation is not described in ConsProb's paper. However, you can easily derive this context profile computation by customizing ConsProb's main inside-outside algorithm for computing posterior nucleotide pair-matching probabilities, as CapR is based on McCaskill's algorithm. The below is examples of ConsProb's average context profiles:
 |
|---|
Docker Playground 
I offer my Docker-based playground for RNA software and its instruction to replay my computational experiments easily.
Method Digest
LocARNA-P can compute posterior nucleotide pair-matching probabilities on RNA pairwise structural alignment. However, LocARNA-P simplifies scoring possible pairwise structural alignments by utilizing posterior nucleotide base-pairing probabilities on RNA secondary structures. In other words, LocARNA-P does not score possible pairwise structural alignments at the same level of scoring complexity as many RNA folding methods. More specifically, many RNA folding methods such as RNAfold score possible RNA secondary structures distinguishing RNA loop structures, whereas many structural alignment-based methods such as LocARNA-P score possible pairwise structural alignments ignoring RNA loop structures. As an antithesis to these structural alignment-based methods, I developed ConsProb implemented in this repository. Distinguishing RNA loop structures, ConsProb rapidly estimates various pairwise posterior probabilities, including posterior nucleotide pair-matching probabilities. ConsProb summarizes these estimated pairwise probabilities as average probabilistic consistency, marginalizing multiple RNA homologs to each RNA homolog.
Author
License
Copyright (c) 2018 Heartsh
Licensed under the MIT license.
Dependencies
~60MB
~1.5M SLoC